研究成果
アジアイネとアフリカイネの雑種障壁を克服した稔性雑種の育成手法を確立
―4倍体化と2倍体化で稔性を持つ中間的遺伝構成雑種を安定育成―
press202515.pdf1.14 MB
令和7年9月25日
国際農研
北海道大学
|
ポイント
|
概要
国際農研と北海道大学の共同研究グループは、アジア栽培イネ (Oryza sativa、以下「アジアイネ」)とアフリカ栽培イネ (Oryza glaberrima、以下「アフリカイネ」) 遺伝子をほぼ均等に持ち、種子稔性を示す種間雑種を効率的に育成する新たな手法を確立しました。
アフリカイネは、病害や環境ストレスへの耐性、雑草競合性など、アジアイネには無い有用特性を備えた栽培種1)であり、育種資源として高く評価されています。しかし両種を交配すると、F1雑種2)はほぼ完全な花粉不稔となり、種子が得られないため、「雑種障壁3)」と呼ばれる大きな育種上の制約が存在していました。この花粉不稔は、雑種不稔遺伝子 (HS遺伝子) 4) の対立遺伝子 (アレル) 5) がヘテロ接合型6) となった場合に発現します。先行研究では、両種のゲノム7) を倍加8) させた4倍体F1雑種において、HS遺伝子の作用が軽減され、花粉稔性が部分的に回復することが明らかになっていました。
本研究では、この知見を基に、4倍体雑種を花粉培養9) して2倍体雑種を作出し、さらにその花粉を培養・倍加して、すべてのHS遺伝子をホモ接合型6) に固定した2倍体の倍加半数体10) 22個体を得ました。そのうち10個体は自家受粉により安定した種子稔性を示し、アジアイネとアフリカイネの双方のゲノムを中間的に保持していました。
この成果は、4倍体化と2段階の花粉培養を組み合わせることで、両種の中間的な遺伝構成を持ちながら種子稔性を確保した種間雑種を安定的に育成できることを初めて実証したものです。今後は、得られた雑種材料の形質評価と有用遺伝子の特定を進め、アジアイネとアフリカイネの長所を併せ持つ新たな育種戦略への展開が期待されます。
本研究成果は、国際科学専門誌「Theoretical and Applied Genetics」オンライン版 (日本時間2025年6月27日) にオープンアクセスで掲載されました。
関連情報
- 予算
- 運営費交付金プロジェクト「熱帯性作物の持続的生産に向けた遺伝資源の情報整備と利用促進技術の開発および国内外との連携強化」
科研費 No. JP19H00937、JP23H02180
発表論文
- 論文著者
- Kuniyoshi, D. and Kishima, Y.
- 論文タイトル
- Fertile interspecific diploid hybrids between the Asian and African rice species facilitated by tetraploidization and its reduction
- 雑誌
- Theoretical and Applied Genetics
DOI: https://doi.org/10.1007/s00122-025-04901-3
問い合わせ先など
国際農研 (茨城県つくば市) 理事長 小山修
- 研究推進責任者:
- プログラムディレクター 飯山みゆき
- 研究担当者:
- 熱帯・島嶼研究拠点 研究員 國吉大地
- 広報担当者:
- 情報広報室長 大森圭祐
プレス用 e-mail : koho-jircas@ml.affrc.go.jp
北海道大学 (北海道札幌市) 総長 寳金清博
- 研究担当者:
- 北海道大学 大学院農学研究院 教授 貴島祐治
- 広報担当者:
- 北海道大学 社会共創部広報課
プレス用 e-mail : jp-press@general.hokudai.ac.jp
研究の背景
イネ (Oryza) 属には、アジアイネとアフリカイネの2種類の栽培種が存在します。現在、世界で栽培されているイネのほとんどはアジアイネであり、高収量や良食味を備えた新品種も、主にアジアイネ同士の交雑により作出されています。
一方、アフリカイネは西アフリカの限られた地域で栽培されており、アジアイネには見られない病害抵抗性や環境ストレス耐性、雑草競合性など有用な特性を備えています。そのため、両者を交雑できれば、これまで栽培が困難であった地域や環境条件でも生育可能な新品種の開発につながると期待されています。
しかし、アジアイネとアフリカイネを交配して得られるF1雑種は、種間の雑種障壁により著しい花粉不稔性11) を示し、自家受粉により種子を実らせることができません。そのため、F1雑種にアジアイネの花粉を受粉させる戻し交雑を繰り返し行い、後代から自家受粉できる系統を選抜することで、アフリカイネをアジアイネの育種に利用してきました。代表例であるネリカ系統12) では、ゲノムの最大1割がアフリカイネ由来であり、アジアイネに由来する高収量性や良食味性に加え、アフリカイネ由来の部分的な栽培特性を併せ持っています。しかしながら、当初期待されていた雑草競合性や低肥沃土壌への適応性といった複雑な形質は十分に継承されていないことが報告されています。したがって、アフリカイネが持つ有望な形質を効果的に育種利用するためには、種間の雑種障壁を克服し、より多くのアフリカイネのゲノムを取り込んだ系統を効率的に育成する手法の開発が不可欠でした。
研究の経緯
アジアイネとアフリカイネはいずれも、2セットのゲノムを持つ「2倍体」の植物です。これまでの研究では、両種の2倍体同士を交配したF1雑種は花粉不稔となる一方、ゲノムを倍加して得た「4倍体」雑種では、花粉稔性が部分的に回復することが示唆されていました。
先行研究 (Kuniyoshi et al., 2024. GENETICS ) では、両種の4倍体同士を交配してF1雑種を作出した結果、2倍体F1雑種では花粉稔性が0%で不稔となる組み合わせでも、4倍体F1雑種では花粉稔性が5.7~28.1%まで回復し、自殖種子が得られることを確認しました (図1)。さらに、この稔性回復は、花粉不稔の原因である雑種不稔遺伝子 (HS遺伝子) の作用が4倍体化により軽減されたためであることも明らかになりました (令和6年度 国際農林水産業研究成果情報)。
また、これまでの多くの先行研究から、アジアイネとアフリカイネの種間雑種における花粉不稔の主因は、複数の雑種不稔遺伝子 (HS遺伝子) であることが分かっていました。HS遺伝子による花粉不稔は、アジアイネ型アレル5) とアフリカイネ型アレルがヘテロ接合型になった場合にのみ発現します。したがって、各HS遺伝子をホモ接合型で固定できれば、種間雑種であっても花粉不稔は生じず、通常の稔性を示す可能性が考えられました。
今回の研究では、この4倍体雑種の倍数性13) を通常のイネのゲノム数である2倍体に戻すことで、両種の中間的な遺伝構成を持ちながら稔性を示す種間雑種の作出が可能であることを実証しました。
研究の内容と成果
- 先ず、4倍体F1雑種の花粉 (染色体数は2倍体相当) を培養することにより、HS遺伝子の一部がホモ接合型に固定された2倍体雑種を作出しました (図2)。得られた2倍体雑種では、ホモ接合型で固定されたHS遺伝子座の数が増えるにつれて花粉稔性が高くなる傾向を示し、最大で26.4%の花粉稔性が確認されました。
- 次に、2倍体雑種の花粉をさらに培養し、HS遺伝子を含む全遺伝子座をホモ接合型に固定した2倍体の倍加半数体 (DH) を22個体獲得しました。そのうち10個体は自家受粉によって安定した種子稔性を示し、花粉稔性は最大92.8%、種子稔性は最大89.8%に達しました。これにより、親系統と同等の高い稔性が得られました (図3)。これら10個体のゲノムを解析した結果、アジアイネ型アレルとアフリカイネ型アレルの構成割合は個体ごとに異なっていました。そのうちの2系統 (DH#124、DH#201) は、両種のゲノムをほぼ同じ割合で受け継いだ中間的な遺伝構成を有することが明らかになりました。
- 以上の結果に基づき、これまで困難とされていたアジアイネとアフリカイネの中間的な遺伝構成を持ちながら稔性を示す種間雑種の育成が、4倍体化とその後の2倍体化の工程を経ることで可能となることが示されました (図4)。
今後の予定・期待
アフリカイネは、アジアイネには無い特性を持つ、もう一つの栽培イネ種であり、アジアイネ育種における有望な遺伝資源として知られていました。しかし、これまで雑種障壁の存在により、その育種利用は限定的で、特に複数の遺伝子が関与する雑草競合性などの複雑な形質をアジアイネへ導入することは困難でした。
今回の研究で確立した稔性雑種の育成手法によって、アジアイネとアフリカイネの遺伝子をほぼ均等に持つ雑種材料を作出できるようになりました。これらの材料は、従来の交雑種であるネリカ系統と比べて、アフリカイネ由来のゲノムを高い割合で保持していることから、アフリカイネが有する雑草競合性、病害や環境ストレスへの耐性などの複雑な有用形質を受け継いでいる可能性があります。
今後は、この雑種材料について農業形質を詳細に評価し、有用形質と関連する遺伝子を特定することで、アフリカイネとアジアイネ双方の長所を併せ持つ新品種の育成に貢献できると期待されます。
用語の解説
- 1) 栽培種
- 人類が長い時間をかけて選抜し、改良してきた植物の品種のことを指します。自然環境に自生している原種は野生種と呼びます。アジアイネとアフリカイネは、アジアおよびアフリカで、それぞれ異なる野生種から選抜され、栽培種となりました。
- 2) F1雑種
異なる2つの品種や種を交配して得られる、第一代 (First Filial、F1) の雑種植物のことです。
- 3) 雑種障壁
- 異なる種 (しゅ) の植物同士を交配した場合に、その雑種の種子が得られなかったり、雑種植物がうまく育たない場合があり、その現象の総称です。
- 4) 雑種不稔遺伝子 (HS遺伝子)
- アジアイネとアフリカイネの雑種は花粉不稔となりますが、この花粉不稔の原因となるのが雑種不稔 (Hybrid Sterility) 遺伝子であり、略してHS遺伝子と呼びます。HS遺伝子は複数存在し、それぞれが独立して花粉不稔を引き起こします。
- 5) 対立遺伝子 (アレル)
- 同じ働きを持つ遺伝子であっても、わずかに機能や構造が異なる「型」が存在します。それぞれの型を「対立遺伝子 (アレル)」と言います。雑種不稔遺伝子には、アジアイネ型のアレルとアフリカイネ型のアレルがあります。
- 6) ヘテロ接合型・ホモ接合型
- 2倍体の植物は、ゲノムを2セット持つため、ある一つの遺伝子について2つのアレルを保持しています。2つのアレルが異なる場合を「ヘテロ接合型」、同じ場合を「ホモ接合型」と言います。雑種不稔遺伝子について、アジアイネはアジアイネ型アレルを、アフリカイネはアフリカイネ型アレルをホモ接合型で保持しています。アジアイネとアフリカイネのF1雑種は、それぞれの親から異なるアレルを一つずつ受け継ぐため、すべての雑種不稔遺伝子がヘテロ接合型になります。
- 7) ゲノム
- 生物が生存するために必要な、一揃いの遺伝子セットのことです。
- 8) 倍加
- 生物が持つゲノムの数を倍にする操作のことです。今回の実験では、イネの分裂中の細胞に対してコルヒチンという物質を作用させることで、ゲノムの倍加を誘導しています。
- 9) 花粉培養
- イネの花粉や、花粉を含む組織 (葯) を培養して、花粉由来のイネ植物体を得る技術です。4倍体イネの花粉は2セットのゲノムを持つため、これを培養すると2倍体のイネが得られます。2倍体イネの花粉を培養すると半数体が得られ、半数体のゲノムを倍加 (2倍に増やすこと) することで、2倍体を得ることができます。
- 10) 倍加半数体
- ゲノムを1セットしか持たない植物を「半数体」と呼びます。このゲノム全体をコピーして2倍体にしたものを「倍加半数体」と言います。倍加半数体は、2つのゲノムセットが完全に同じであるため、すべての遺伝子がホモ接合型になっています。
- 11) 不稔性
- 花粉や種子が正常に発育できず、子孫を残せない状態を指します。
- 12) ネリカ (NERICA、New Rice for Africa) 系統
- アジアイネとアフリカイネの雑種である陸稲品種群です。18系統あり、代表品種であるネリカ4は、ゲノムの約1.3%がアフリカイネに由来しています。
- 13) 倍数性
- ゲノムのセット数のことを指します。ゲノムを2セット持つ植物は「2倍体」と呼ばれ、イネを含む多くの植物はこの2倍体の状態です。イネにコルヒチンを作用させることで、ゲノムを4セット持つ「4倍体」を作成できます。逆に、2倍体の花粉を培養することで、ゲノムが1セットだけの「半数体」を作成することも可能です。
担当研究者の声

熱帯・島嶼研究拠点
研究員 國吉大地
アジアイネとアフリカイネ種間の稔性雑種の育成手法に取り組み始めたのは学生の頃 (2013年) でした。10年以上かかりましたが、一つの道筋を示すことができてよかったです。
今後は雑種系統が両種の特性をどのように引き継いでいるのかを明らかにしていきたいです。アフリカイネを含めた近縁イネ種を利用した雑種品種をサブサハラ・アフリカでリリースするのが私の夢です。
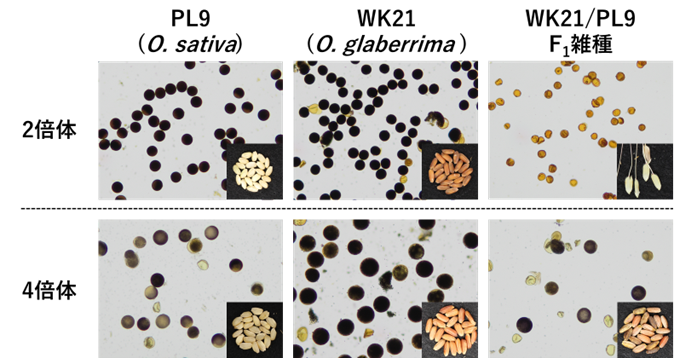
図1 親系統およびF1雑種の花粉稔性
開花期の花粉をヨウ素液で染色し、顕微鏡で観察しました。黒く染まった丸い花粉は稔性を持っています。親系統であるPL9 (Oryza sativa) とWK21 (Oryza glaberrima) の花粉はほぼすべて、黒く丸い形をしています。2倍体のF1雑種では、すべての花粉が不稔性でしたが、4倍体F1雑種では花粉稔性が約30%まで回復しました。
各写真の右下には、それぞれの植物材料から得られた自殖種子を示しています。2倍体F1雑種は花粉不稔のため自殖種子が得られませんでしたが、花粉稔性が回復した4倍体F1雑種からは、十分な量の自殖種子を得ることができました。
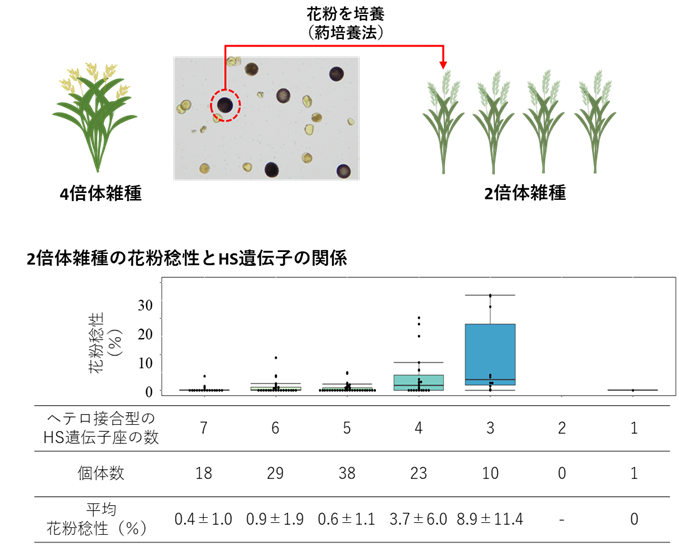
図2 4倍体雑種の花粉培養で獲得した2倍体雑種の花粉稔性
花粉は、親植物の半数セットのゲノムを保持しているため、4倍体植物の花粉は2倍体相当のゲノムを保持しています。そこで、4倍体雑種の花粉を培養して2倍体系の雑種植物を作出し、7つのHS遺伝子座の接合型 (ホモ接合またはヘテロ接合) と花粉稔性との相関を確認しました。2倍体F1雑種では、7つのHS遺伝子座がヘテロ接合型で保持されていますが、4倍体の花粉に由来する2倍体雑種では、ヘテロ接合型のHS遺伝子座の数が7から1まで様々であり、ヘテロ接合型HS遺伝子座の数が減少するにつれて花粉稔性が高くなる傾向が観察されました。ヘテロ接合型HS遺伝子座が1あるいは2の植物は、統計的に得られる確率が低く、今回の試験では1個体のみが得られましたが花粉不稔でした。

図3 2倍体倍加半数体について
2倍体雑種の花粉 (半数体) を培養し、さらにゲノムを倍加させることで、すべてのHS遺伝子がホモ接合型で固定された2倍体の倍加半数体 (DH) を作出しました。全22個体の DH植物のうち、10個体 (図示) は自家受粉により安定した種子稔性を示しました。
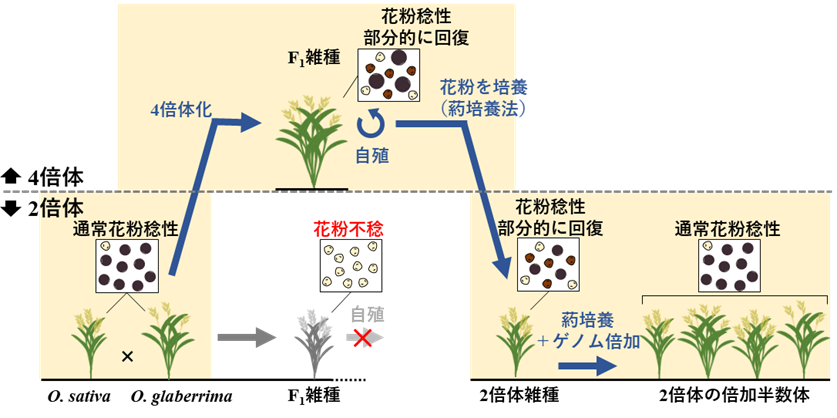
図4 4倍体化と2倍体化を介した稔性雑種の育成手法
アジアイネとアフリカイネ種間の雑種は、雑種障壁により種子が実りません。しかし、雑種障壁を生み出しているHS遺伝子の効果を4倍体化によって一時的に回避し、その後もう一度2倍体に戻すことで、両種の中間的な遺伝構成を持つ2倍体の稔性雑種を育成できることが明らかになりました。