イネ複数品種におけるゲノム編集系および効率的な変異系統獲得手法
要約
未熟胚を用いたアグロバクテリウム法は、主要イネ品種へのCRISPR/Cas9システム導入によるゲノム編集に有効である。Cas9遺伝子が除去された形質転換後代の個体を選抜することで、キメラ性および分離異常を示さない変異系統を効率的かつ確実に作出できる。
背景・ねらい
CRISPR/Cas9(クリスパー・キャスナイン)は標的遺伝子に変異を誘起するゲノム編集技術の一つであり、変異導入による作物の農業形質改良への利用が期待されている。これまでのイネにおけるCRISPR/Cas9に関する報告で用いられた品種は、形質転換効率が高い日本晴などに限られており、CRISPR/Cas9をイネの分子育種に応用するためには、各地の主要品種に適用可能な系を確立する必要がある。また、CRISPR/Cas9により誘起したイネの変異遺伝子はメンデルの法則に従って分離するとされているが、変異遺伝子の分離および後代におけるCRISPR/Cas9の機能についての検討は不十分であり、変異遺伝子をホモ接合型で保持する個体を効率的に作出するためには、これらを明らかにする必要がある。本研究では、各地の主要品種を含むイネ5品種におけるCRISPR/Cas9によるゲノム編集系の確立を試みるとともに、後代におけるCas9遺伝子と標的遺伝子の分離との関係性を明らかにし、効率的かつ確実に変異型ホモ個体を獲得する方法を確立する。
成果の内容・特徴
- 未熟胚を用いたアグロバクテリウム法は、日本型イネ品種日本晴、コシヒカリ、NERICA1およびCuringa、およびインド型イネ品種IR 64へのCRISPR/Cas9システムの導入に有効である。得られる形質転換体の大部分は標的遺伝子の変異体である(表1)。
- 単一対立遺伝子変異体の形質転換当代 (T0) の自殖後代 (T1) においては、変異体の出現頻度がメンデルの法則に基づく推定値よりも高くなる分離異常が多く見られる。また、両対立遺伝子変異の細胞および単一対立遺伝子変異の細胞両方を有するキメラ個体が出現する場合がある(図1)。
- 単一対立遺伝子変異体のT1がCas9遺伝子を保持する場合、その後代のT2においても分離異常およびキメラ個体の出現が認められる。一方、T1がCas9遺伝子を保持しない場合、T2における変異遺伝子の分離はメンデルの法則に従い、キメラ個体は出現しない。このことは、後代においてCas9遺伝子は新規変異を誘起し、後代個体のキメラ性および分離異常の要因となることを示す(図2)。
- Cas9遺伝子を保持しないT1個体を選抜することにより、分離異常およびキメラ個体出現の可能性が排除されるため、効率的かつ確実に変異型ホモ個体を獲得することができる。
成果の活用面・留意点
- 特定の遺伝子変異による形質が育種上有用である場合、本成果を用いることで各地の主要品種に有用形質を付与することができる。
- 一般的にCRISPR/Cas9によるゲノム編集において生じる可能性があるT0の体細胞変異に伴う分離異常、標的遺伝子以外の変異、および培養変異等にも留意する必要がある。
- 変異体の実用化に際しては、知的財産権の取扱いに留意する必要がある。
具体的データ
-
表1 未熟胚を用いたアグロバクテリウム法を用いてイネ品種に導入したCRISPR/Cas9システムによる効率的な標的遺伝子の変異誘起
品種形質転換体の数単一対立遺伝子変異体の数両対立遺伝子変異体の数変異体の数の合計日本晴106266490コシヒカリ1711011NERICA154163349Curinga27213780217IR645415計454184188372標的遺伝子はカロテノイド合成のキー酵素であるphytoene dasaturase(PDS)遺伝子であり、両対立遺伝子変異体はアルビノとなる。
-
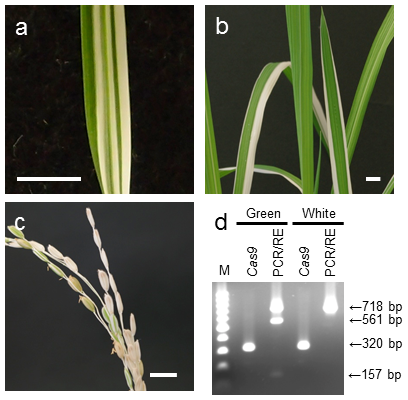
図1 T1におけるキメラ個体の出現
(a)キメラ個体の幼苗の葉。(b)温室で育成したキメラ個体の葉。(c)キメラ個体の穂。バーは1 cm。(d)キメラ個体の緑および白の部分を別々にPDS遺伝子のPCR/RE検定およびCas9遺伝子のPCR検定に供試した。PCR/REの結果、バンドが3本の場合は単一対立遺伝子変異体であり、1本の場合は両対立遺伝子変異体である。 -
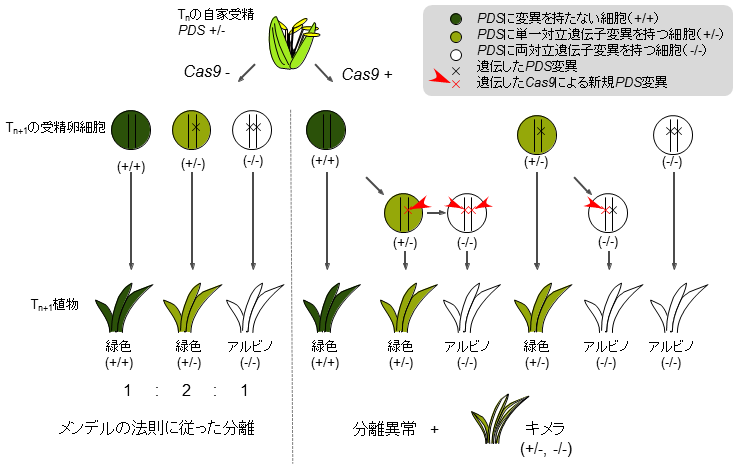
図2 CRISPR/Cas9により導入した変異遺伝子の分離異常発生の模式図
植物がCas9遺伝子を保持しない場合、変異遺伝子はメンデルの法則に従って分離する。植物がCas9遺伝子を保持する場合、Cas9は新規変異を誘起し、分離異常およびキメラ個体発生の要因となる。
- Affiliation
-
国際農研 熱帯・島嶼研究拠点
- 分類
-
研究
- 研究プロジェクト
- プログラム名
- 予算区分
-
交付金 » 不良環境耐性作物開発
- 研究期間
-
2016年度(2014~2016年度)
- 研究担当者
-
石崎 琢磨 ( 熱帯・島嶼研究拠点 )
- ほか
- 発表論文等
-
https://doi.org/10.1007/s11032-016-0591-7
Ishizaki T (2016) Molecular Breeding, 36
- 日本語PDF
-
A4 381.14 KB
A3 311.62 KB
- English PDF
-
A4 664.3 KB
A3 619.81 KB
- ポスターPDF
-
Poster 419.24 KB