Selective logging criteria to ensure healthy seed production for dipterocarp species that depend their pollination on strong flyer insects
Description
Timber in Malaysia has been produced from tropical rainforests that were recognized as one of the highest biodiversity hotspots in the world. Trees of timber species larger than 50 cm in trunk diameter at breast height (dbh) have been harvested in ongoing selective logging operation in Malaysia. The harvesting lowered remaining adult tree density, which might inhibit pollen travel from a tree to others by pollinator insects. This is a critical issue to produce healthy seeds that shall be source of forest regeneration, because it has been reported that mother trees at lowered population density show frequent self-fertilization that produces less vigorous seeds. A group in genus Shorea known as ‘red meranti’ in forestry trading name is belonging to some sections, Mutica, Brachypterae etc., which is most abundant in lowland and hill dipterocarp forests. These species produce small-sized flower, which is for symbiosis with thrips, main pollen vector of these species, characterized by weak flying ability. The reproductive system ensured by pollination of thrips is vulnerable to reduction of tree density, such as the selective logging activity.
On the other hand, recent studies on pollen dispersal pattern of dipterocarps revealed that pollen dispersal pattern was mainly regulated by pollinator and conspecific tree density. Another group in genus Shorea known as ‘balau’ in forestry trading name is belonging to section Shorea, which produces the second highest valuable timber in Peninsular Malaysia. The species belonging to this section produces relatively larger flower than ‘red meranti’ species, which putatively attract small beetles belonging to Chrysomelidae. The small beetles are generally characterized as energetic strong flyer, which should represent different pollen dispersal pattern from ‘red meranti’ with thrips pollination. We estimated pollen dispersal pattern by using paternity analysis of seeds and a reproductive model (JIRCAS Research Highlight No. 15, 2011), and evaluated selective logging criteria by simulating pollen dispersal before and after selective logging for S. maxwelliana, one of the ‘balau’ species.
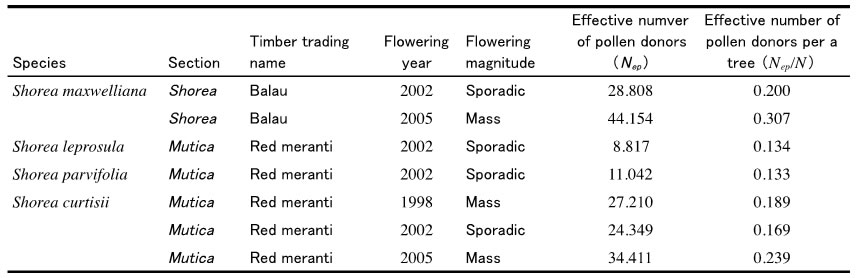
When the pollen dispersal pattern was compared between mass flowering season (in 2005) and sporadic flowering season (in 2002), the strong flying ability of small beetles achieved active pollen dispersal even though flowering tree density was reduced in the sporadic flowering season (Fig. 1). However, seeds with higher genetic diversity were produced in the mass flowering season, because more flowering trees in the mass flowering seasons contributed as pollen donors, which was shown by number of effective pollen donors (Nep) in Table 1. We calculated rate of outcrossing pollen over the mother trees in after the logging to in before logging. The criterion of selective logging was increased with 1 cm from 40 cm to 100 cm at dbh in the simulation. Only more than about trees with 80 cm dbh can be harvested to conserve 50% of outcrossing pollen over the mother trees in S. curtisii (one of the ‘red meranti’ species). However, our result showed that trees with more than about 60 cm dbh can be harvested for ‘balau’ because of the small beetles’ stronger flying ability (Fig. 2).
Our results showed that the selective logging criterion should be determined for every timber trading group of dipterocarps because reproductive characteristics within the timber trading group were similar. Our reproductive model and the simulation assumed similarity of pollinator and conspecific tree density. When the results would be applied to different type of forests, the simulation results couldn’t be directly applicable to the practices.
Figure, table
-
Table 1. The comparison of number of effective pollen donors between 'balau' and 'red meranti'
-
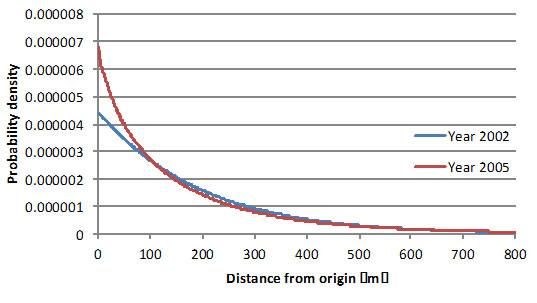
Fig. 1. Pollen dispersal pattern of Shorea maxwelliana recognized as ‘balau’ in two flowering events with different flowering maganitude (left).
-
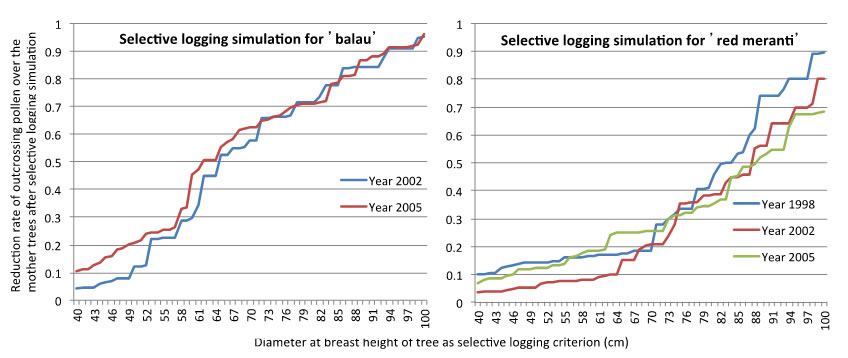
Fig. 2. Reduction rate of outcrossing pollen over the mother trees after selective logging simulation with every 1cm increment of cutting limit from 40 to 100cm (below).
- Affiliation
-
Japan International Research Center for Agricultural Sciences Forestry Division
- Classification
-
Administration B
- Program name
- Term of research
-
FY 2013 (FY 2006-FY 2015)
- Responsible researcher
-
Tani Naoki ( Forestry Division )
ORCID ID0000-0001-9878-6352KAKEN Researcher No.: 90343798Tsumura Yoshihiko ( Forestry and Forest Products Research Institute )
KAKEN Researcher No.: 20353774Muhammad Norwati ( Forest Research Institute Malaysia )
Lee Soon Leong ( Forest Research Institute Malaysia )
- ほか
- Publication, etc.
-
https://doi.org/10.1371/journal.pone.0082039
Masuda, S. et al. (2013) PLoS ONE 8(12): e82039
Tani, N. et al. (2012) JIRCAS Working Report No.76: 61-66
- Japanese PDF
-
2013_C09_A3_ja.pdf258.92 KB
2013_C09_A4_ja.pdf413.83 KB
- English PDF
-
2013_C09_A3_en.pdf91.4 KB
2013_C09_A4_en.pdf601.88 KB
- Poster PDF
-
2013_C09_poster.pdf277.84 KB