Enhancement of porosity and aerenchyma formation by nitrogen deficiency in rice roots (Oryza sativa L.)
Description
Lysigenous aerenchyma is formed by cell collapse accompanied with cell death. The aerenchyma in roots is concerned in water logging tolerance of plants by providing oxygen from ground portion to roots. Field crops such as wheat show generally poor tolerance to water logging, while semiaquatic crops such as rice show high tolerance to water logging. One of the major reasons is thought to be caused by difference of initiation in aerenchyma formation between these crops. In wheat, aerenchyma is inducibly formed by trigger of multiple environmental factors such as oxygen and nutrients-nitrogen, phosphorus and potassium-deficiency. While, rice can form two kinds of aerenchyma, constitutive and inducible aerenchyma, respectively. Thus, mechanism of aerenchyma formation in rice is more complicated than that in field crops. Moreover, it is still unknown mechanism of aerenchyma formation induced by nitrogen deficiency, although the resulting aerenchyma is likely to benefits reduction of energy loss. In this study, we attempted to (1) establish reliable growth condition to estimate aerenchyma formation and (2) reveal pattern of aerenchyma formation induced by nitrogen deficiency in rice roots.
Before evaluating aerenchyma formation, we modified growth conditions, e.g., hydroponic solution and growth period, to estimate precisely response by nitrogen deficiency alone. Finally, we could establish precise growth condition demonstrating recovery of growth vigor caused by pH reduction in hydroponic solutions (Fig. 1). As compared with nitrogen sufficient, nitrogen deficiency facilitated formation of air space of roots demonstrating increase of porosity which is air space on whole roots (Fig. 2). In order to determine spatial and temporal pattern of aerenchyma formation induced by nitrogen deficiency, cross sections of seminal root of seedlings grown nitrogen deficiency alone and oxygen deficiency alone were prepared at positions from root tip to base continuously. Microscope observations revealed that aerenchyma formation was enhanced in both nitrogen and oxygen deficiency compared with reference condition (Fig. 3). In nitrogen deficiency, aerenchyma formation initiated at position close to root base. Conversely, the initiation was observed at position close to root tip (Fig. 3).
As far as we know, this is first evidence for enhancement of porosity and aerenchyma formation by nitrogen deficiency in rice roots. It strongly indicates that physiological role of induced aerenchyma by nitrogen deficiency distinct from that by oxygen deficiency, demonstrating the different initiation pattern of aerenchyma between nitrogen and oxygen deficiency. Aerenchyma induced by nitrogen deficiency may function in reducing respiration and remobilization of nitrogen, or both. Furthermore, our growth condition established is expected to isolate casual genes associated with aerenchyma formed constitutively and inducible toward developing molecular breeding techniques for conferring tolerant to water logging in field crops in near future.
Figure, table
-
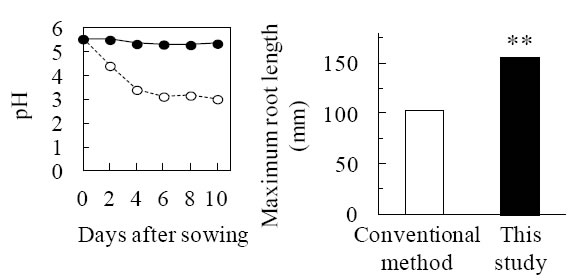
Fig.1. pH maintenance of nutrient solutions (left) and recovery of root elongation (right) in this study (improved method).
○: conventional method, ●: this study Asterisks (**) represent a significant difference in maximum root length between conventional method and improved method at P-value of 1% level (one-way ANOVA). -
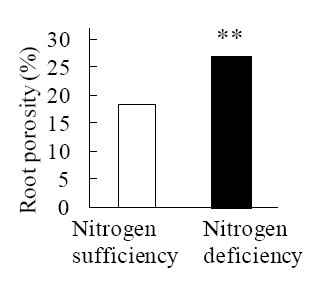
Fig. 2. Increased root porosity by nitrogen deficiency.
Root porosity is size of air space including aerenchyma. Ten-days-old seedlings were used. Asterisks (**) represent a significant difference in root porosity between nitrogen sufficient condition and nitrogen deficient condition at P-value of 1% level (one-way ANOVA). -
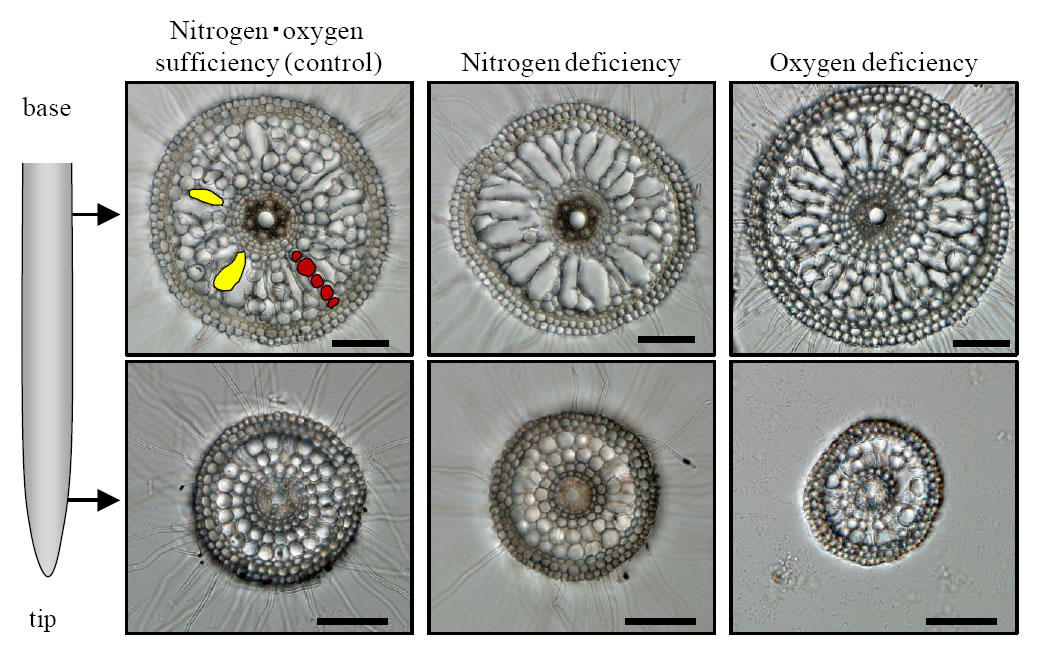
Fig. 3. Root aerenchyma in rice.
Increased aerenchyma was formed by nitrogen deficiency or oxygen deficiency. Examples of cortical cell, which are living cells were illustrated with red. Examples of aerenchyma, which are dead cells were illustrated with yellow. Scale bar in individual pictures indicates 100 µm.
- Affiliation
-
Japan International Research Center for Agricultural Sciences Biological Resources and Post-harvest Division
- Classification
-
Administration A
- Research project
- Program name
- Term of research
-
FY 2013 (FY 2011-FY 2015)
- Responsible researcher
-
Obara Mitsuhiro ( Biological Resources and Post-harvest Division )
Abiko Tomomi ( Biological Resources and Post-harvest Division )
- ほか
- Publication, etc.
-
https://doi.org/10.1016/j.plantsci.2013.10.016
Abiko,T. and Obara, M. (2014) Plant Science, 215-216: 76-83
- Japanese PDF
-
2013_B02_A3_ja.pdf712.42 KB
2013_B02_A4_ja.pdf322.88 KB
- English PDF
-
2013_B02_A3_en.pdf204.89 KB
2013_B02_A4_en.pdf513.67 KB
- Poster PDF
-
2013_B02_poster.pdf399.18 KB